
Musicians have more robust and efficient neural responses in the cortical and sub-cortical regions, demonstrating that musical experience benefits the processing of both non-linguistic and linguistic stimuli.
ObjectiveThis study aimed to verify P300's latency and amplitude behavioral using contralateral stimulation in musicians and non-musicians.
MethodsThis was a case–control study. Subjects were divided in two groups: musicians, comprising 30 professional musicians, and non-musicians, comprising 25 subjects without musical experience.
ResultsThe present study showed that the musicians had lower latencies and higher amplitudes than the non-musicians in the P300 without contralateral noise. For the P300 amplitude values, the difference between groups persisted, and the musicians presented significantly higher amplitude values compared with the non-musicians; additionally, the analysis of the noise effect on the P300 response showed that the latency values were significantly increased in the musicians.
ConclusionThe central auditory nervous system of musicians presents peculiar characteristics of electrophysiological responses probably due to the plasticity imposed by musical practice.
Os músicos possuem respostas neurais mais robustas e eficientes em regiões corticais, mostrando que a experiência musical beneficia o processamento de estímulos linguísticos e não linguísticos.
ObjetivoVerificar como a latência e a amplitude do P300 se comporta usando estimulação contralateral, em músicos e não músicos.
MétodoEstudo de caso-controle. Os indivíduos foram divididos em dois grupos: GM (grupo de músicos) com 30 músicos profissionais e GNM (grupo de não músicos) com 25 indivíduos sem experiência musical.
ResultadosOs resultados mostraram que: GM teve latências menores e amplitudes maiores do que a GNM no P300 sem ruído contralateral. Para os valores de amplitude do P300, a diferença entre os grupos se manteve, e o GM apresentou valores de amplitude significativamente maiores em comparação com o GNM; e a análise do efeito do ruído sobre a resposta P300 mostrou que os valores de latência foram significativamente maiores no GM.
ConclusãoConcluímos que o sistema nervoso auditivo central de músicos apresenta características peculiares de respostas eletrofisiológicas provavelmente devido à plasticidade imposta pela prática musical.
The effects of musical experience on the representation of sounds in the auditory cortex have been functionally and anatomically observed in several studies. Anatomical differences were found in various brain structures, such as the Heschl's gyrus, the secondary auditory cortex, the corpus callosum, and the temporal plane of musicians.1
A study by Ohnish et al. in 2001, using functional magnetic resonance imaging (FMRI), demonstrated that passive music listening produces significant activation in the superior temporal and bilateral medial gyrus, together known as the auditory association cortex, in both musicians and non-musicians. Musicians show right dominant temporal cortical activation, whereas non-musicians show left dominant temporal cortical activation.2
Chermak stated that, by stimulating different areas of the brain, such as the frontal, temporal, parietal, and sub-cortical regions, music can help improve several attention-related functions, including memory, learning, language, and even emotional aspects.3
Several authors also agree that musicians have more robust and efficient neural responses in the cortical and sub-cortical regions, demonstrating that musical experience benefits the processing of both non-linguistic and linguistic stimuli. Thus, musical training has been a frequent and effective resource in the rehabilitation of communication disorders.3–5
Auditory evoked potentials (AEPs) are important tools in the investigation of auditory function because, in addition to being objective and noninvasive tests, they are sensitive to plastic changes in the auditory pathway that result from external stimulation.6
P300 is an endogenous long-latency auditory evoked potential (LLAEP) that depends on the conscious response of the individual and appears as a positive wave between 50 and 500ms after acoustic stimulus presentation, with amplitude ranging from 7 to 25μV. This potential is elicited in an auditory discrimination task termed “the oddball paradigm”, and is also known as a cognitive potential as it is used to investigate cognitive abilities, such as discrimination and attention. The P300 generators are still being investigated, but it is known that P3 is originated by different structures not only in the cortex, but also in sub-cortical regions. Thus, it can be said that P300 comprises responses from frontal and centro-parietal cortex, as well as the hippocampus.7 There are evidences that subthalamus and medial geniculate body contributes to P300 generation, with some activities at the orbital gyrus, rostral thalamus, and anterior commissure.8,9 Moreover, the current investigations regarding late potentials focus on the relationship between their characteristics and information processing, such as codification, memory, and decision-making.10
AEP studies have shown that the auditory cortex responds to sounds differently in musicians and non-musicians,11,12 and that the auditory cortex plasticity, in relation to the effects of musical training, is higher when the training begins in childhood.2,13 In 2003, Trainor et al. observed improvement in LLAEPs after auditory musical training in both children and adults, showing that music can alter the auditory cortical representation.13 Other authors suggest that this type of stimulation can be used as a tool in auditory rehabilitation.3
In 2006, Lopez et al. observed significant differences in the latency and amplitude values of the potentials mismatch negativity (MMN) and P300 between a group of amateur musicians and a group of non-musicians (NMG); the group of musicians (MG) presented reduced latencies and greater amplitudes.11
Musacchia et al., in 2007, evaluated the brainstem AEPs in a MG, and observed that they exhibited reduced latencies and larger amplitudes when compared with a NMG, both for musical and speech stimuli.5
Even beyond sub-cortical and cortical functional enhancements, musical training may shape auditory function in structures that are as peripheral as the cochlea; musicians demonstrated a greater degree of efferent control over outer hair cell activity along the basilar membrane than non-musicians.14,15 Such comprehensive perceptual and neural enhancements may be driven, at least partially, by strengthened cognitive control over basic auditory processing, as engendered by auditory attention,16–18 two auditory cognitive skills that are enhanced in musicians.
Krishnamurti, in 2001, observed the influence of contralateral competitive noise in P300 latency and amplitude. According to the author, the introduction of contralateral competitive noise complicates the situation, increasing the difficulty of P300 discrimination and consequently resulting in increased latency and decreased amplitude of this potential.19
Although behavioral studies have found similar effects in musicians and non-musicians,20 the results with event-related potentials (ERPs) are inconsistent.21
Previous studies have demonstrated the reduction of cochlear response – decreased amplitude and increased latencies – due to inhibitory effect induced by the presence of noise. These changes in responses were observed throughout the auditory pathway, through assessments from the cochlea up to the auditory cortex,22 demonstrating that the auditory system of healthy individuals suffers inhibition effects in the presence of a competitive stimulus.
Moreover, little is known about the physiological mechanisms that underlie the functioning of the efferent pathway. To investigate the positive effect of musical experience on the auditory pathway, this study aimed to assess the latency and amplitude of P300 cognitive potential responses in the presence and absence of contralateral noise in musicians and non-musicians. This study addressed the following research questions: Are there differences in the latencies and amplitudes of P300 auditory ERPs, with and without contralateral noise, between musicians and non-musicians? Can contralateral noise inhibit P300 response in both groups?
MethodsThe present study was conducted at the authors’ lab and was approved by the ethics committee (No. 0874/07).
ParticipantsThis was a case–control study that included 30 professional musicians of both genders who had formally studied music since childhood or teenage years and still practice some kind of musical instrument, between 20 and 53 years of age, who were referred to as the MG. In the control group, 25 subjects of both genders without any formal musical experience, between 18 and 30 years of age, were selected and were referred to as the NMG. All participants signed an informed consent.
As inclusion criteria, it was established that all of the participants should have hearing thresholds within the normal limits at frequencies of 250–8000Hz,23 which was verified through pure tone audiometry, and an absence of hearing complaints and/or neurological disorders. The difference between the groups was determined based on years of musical experience.
The procedures performed in the NMG and MG included the following: P300 with and without contralateral noise. P300 was elicited through the Bio-Logic Traveler Express Auditory Evoked Potential Machine (ANSI, 1996), using TDH 39 earphones. To evaluate P300, the active electrode was positioned at Cz, the reference electrodes were positioned on the right (A2) and left (A1) mastoids (a linked mastoid electrode regardless of ear of presentation), and the ground electrode was positioned at Fz.24 A tone burst at 1500Hz was used as deviant (rare stimulus), presented randomly with 20% probability, mixed with a frequent tone burst stimulus at 1000Hz and presented with 80% probability using the oddball paradigm, 70dB HL intensity and a rate of one stimulus per second. Individuals were asked to verbally count the rare stimuli. P300 was identified as the positive deflection 250–500ms after the stimulus.
To verify the suppression effect, the test was repeated with the use of white noise (WN) in the contralateral ear with the same intensity as the tone burst (70dB HL). The WN was generated by Bio-Logic System. Suppression values for both the latency and amplitude parameters were calculated by subtracting the value obtained with noise from the value obtained without noise (latency value without noise−latency value with noise). The response suppression effect of latency is inversely proportional to the amplitude; thus, while the latency values are expected to increase in the presence of contralateral noise, the amplitude values should decrease.
In the present study, the examiner applying the test was also responsible for the analysis of its results; thus, this was not a blind study. Although it could be argued that such analysis may be influenced by its subjective aspects, the authors do not believe this influence undermines the obtained results. When interference was observed during the sweeping, a new sweep was performed.
The intra- and inter-group analyses were based on the verification of the numerical values of the latency, in ms, and of the amplitudes, in μV, of the P300 in the evaluation with and without contralateral noise and on the comparison of the values obtained in each situation. These data were subjected to the Kolmogorov–Smirnov test, and a normal distribution was observed for all of the datasets.
Subsequently, in a comparison of the mean latency and amplitude in both groups and in both ears, analysis of variance (ANOVA) was applied to determine whether there were significant differences in the latencies and amplitudes of P300 between the MG and the NMG. Student's t-test for dependent samples was used to compare the means obtained by the right and left ears in each group. For intra-group analysis for the effect of contralateral noise on the latencies and amplitudes of P300 within each group, ANOVA with repeated measures was performed. A significance level equal to 0.05 (5%) was adopted. Statistically significant values were marked with an asterisk when ≤0.05. Statistical analysis was performed using the SPSS software.
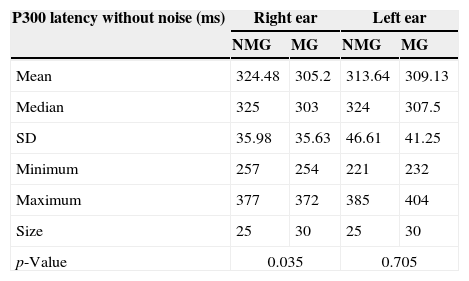
ResultsTable 1 shows descriptive statistics of the data obtained in the comparison of the P300 latency values without noise between the NMG and MG for the right and left ear.
Comparison of the P300 latency values (ms) without contralateral noise between the NMG and MG for the right and left ear.
| P300 latency without noise (ms) | Right ear | Left ear | ||
|---|---|---|---|---|
| NMG | MG | NMG | MG | |
| Mean | 324.48 | 305.2 | 313.64 | 309.13 |
| Median | 325 | 303 | 324 | 307.5 |
| SD | 35.98 | 35.63 | 46.61 | 41.25 |
| Minimum | 257 | 254 | 221 | 232 |
| Maximum | 377 | 372 | 385 | 404 |
| Size | 25 | 30 | 25 | 30 |
| p-Value | 0.035 | 0.705 | ||
NMG, group of non-musicians; MG, group of musicians.
It was observed that the mean P300 latency without contralateral noise in the NMG was greater than that observed in the MG. However, this difference was only statistically significant for the right ear [F1.53=4.67; p=0.035].
Using the t-test for dependent samples, no statistically significant differences were observed in either group in the comparison between the left and right ears for the P300 latency values without contralateral noise [NMG: t(24)=1.052; p=0.30; MG: t(29)=−0.831; p=0.41].
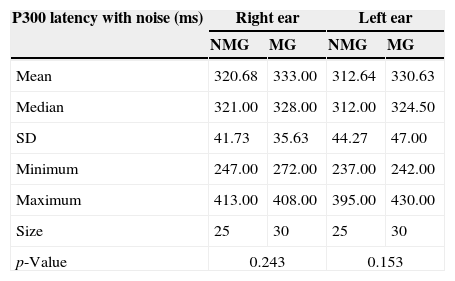
Table 2 presents descriptive statistics of the data obtained in the comparison between the P300 latency values with contralateral noise that were obtained by the NMG and MG for the right and left ears.
Comparison of the P300 latency values (ms) obtained with contralateral noise between the NMG and the MG for the right and left ears.
| P300 latency with noise (ms) | Right ear | Left ear | ||
|---|---|---|---|---|
| NMG | MG | NMG | MG | |
| Mean | 320.68 | 333.00 | 312.64 | 330.63 |
| Median | 321.00 | 328.00 | 312.00 | 324.50 |
| SD | 41.73 | 35.63 | 44.27 | 47.00 |
| Minimum | 247.00 | 272.00 | 237.00 | 242.00 |
| Maximum | 413.00 | 408.00 | 395.00 | 430.00 |
| Size | 25 | 30 | 25 | 30 |
| p-Value | 0.243 | 0.153 | ||
NMG, group of non-musicians; MG, group of musicians.
No statistically significant differences were observed in the comparison of the P300 latency values with contralateral noise between the NMG and the MG for the right and left ears.
No statistically significant differences were observed between the ears in either group for the P300 latency values with contralateral noise [NMG: t(24)=1.100; p=0.28; MG: t(29)=0.425; p=0.67].
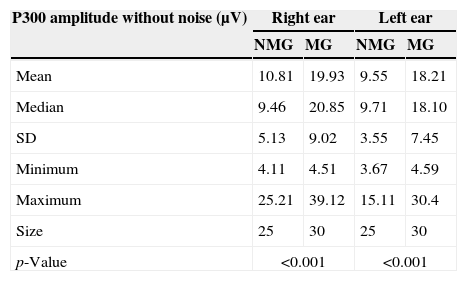
Table 3 shows descriptive statistics of the data obtained in the comparison of the P300 amplitude values without competitive noise between the NMG and the MG for the right and left ears.
Comparison between the P300 amplitude (μV) without contralateral noise between the NMG and the MG for the right and left ears.
| P300 amplitude without noise (μV) | Right ear | Left ear | ||
|---|---|---|---|---|
| NMG | MG | NMG | MG | |
| Mean | 10.81 | 19.93 | 9.55 | 18.21 |
| Median | 9.46 | 20.85 | 9.71 | 18.10 |
| SD | 5.13 | 9.02 | 3.55 | 7.45 |
| Minimum | 4.11 | 4.51 | 3.67 | 4.59 |
| Maximum | 25.21 | 39.12 | 15.11 | 30.4 |
| Size | 25 | 30 | 25 | 30 |
| p-Value | <0.001 | <0.001 | ||
NMG, group of non-musicians; MG, group of musicians.
Statistically significant differences were observed when comparing the P300 amplitude values between the NMG and MG for both the right ear [F1.53=20.06; p<0.001] and the left ear [F1.53=28.25; p<0.0001].
No statistically significant differences were observed in either group in the comparison of the P300 amplitude values between the ears without contralateral noise [NMG: t(24)=1.485; p=0.15; MG: t(29)=1.847; p=0.07].
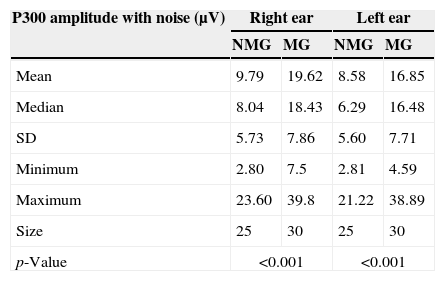
Table 4 shows the comparisons of the NMG and MG with respect to the results found for the P300 amplitude values with contralateral noise for the right and left ear.
Comparison of the P300 amplitudes (μV) with contralateral noise between the NMG and MG for the right and left ears.
| P300 amplitude with noise (μV) | Right ear | Left ear | ||
|---|---|---|---|---|
| NMG | MG | NMG | MG | |
| Mean | 9.79 | 19.62 | 8.58 | 16.85 |
| Median | 8.04 | 18.43 | 6.29 | 16.48 |
| SD | 5.73 | 7.86 | 5.60 | 7.71 |
| Minimum | 2.80 | 7.5 | 2.81 | 4.59 |
| Maximum | 23.60 | 39.8 | 21.22 | 38.89 |
| Size | 25 | 30 | 25 | 30 |
| p-Value | <0.001 | <0.001 | ||
NMG, group of non-musicians; MG, group of musicians.
Statistically significant differences were found in the comparison of the P300 amplitude values with contralateral noise between the NMG and the MG for both the right ear [F1.53=27.03; p<0.001] and the left ear [F1.53=19.902; p<0.001].
In the comparison between the ears for the NMG, no statistically significant differences were observed in the P300 amplitude values with contralateral noise [NMG; t(24)=1.300; p=0.290]. For the MG, a statistically significant difference was found between the right and left ears for the P300 amplitude values with contralateral noise [t(29)=3.357; p=0.002].
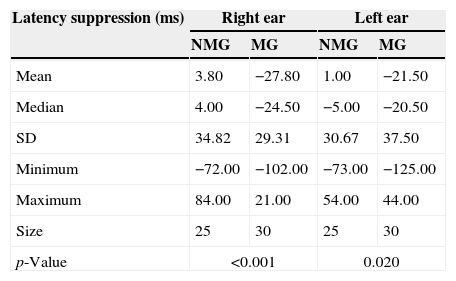
Table 5 shows descriptive statistics for the comparisons between the NMG and the MG regarding the results observed for the P300 latency suppression values.
Comparison of the P300 latency suppression (ms) between the NMG and the MG for the right and left ears.
| Latency suppression (ms) | Right ear | Left ear | ||
|---|---|---|---|---|
| NMG | MG | NMG | MG | |
| Mean | 3.80 | −27.80 | 1.00 | −21.50 |
| Median | 4.00 | −24.50 | −5.00 | −20.50 |
| SD | 34.82 | 29.31 | 30.67 | 37.50 |
| Minimum | −72.00 | −102.00 | −73.00 | −125.00 |
| Maximum | 84.00 | 21.00 | 54.00 | 44.00 |
| Size | 25 | 30 | 25 | 30 |
| p-Value | <0.001 | 0.020 | ||
NMG, group of non-musicians; MG, group of musicians.
Statistically significant differences were found in the comparison of the P300 latency suppression between the NMG and the MG for both the right ear [F1.53=13m35; p=0.001] and the left ear [F1.53=5.77; p=0.020].
No statistically significant differences in the P300 latency suppression values were observed in either group in the comparison between ears [NMG: t(24)=0.308; p=0.76; MG: t(29)=0.950; p=0.35].
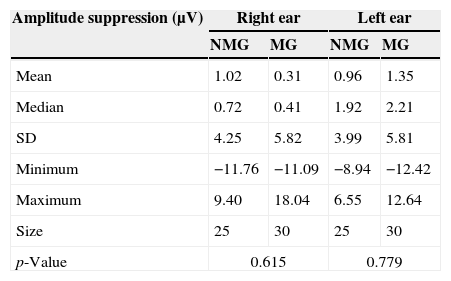
Table 6 presents descriptive statistics for the comparisons between the NMG and the MG regarding the results observed for the P300 amplitude suppression values.
Comparison of the P300 amplitude suppression (μV) between the NMG and the MG for the right and left ears.
| Amplitude suppression (μV) | Right ear | Left ear | ||
|---|---|---|---|---|
| NMG | MG | NMG | MG | |
| Mean | 1.02 | 0.31 | 0.96 | 1.35 |
| Median | 0.72 | 0.41 | 1.92 | 2.21 |
| SD | 4.25 | 5.82 | 3.99 | 5.81 |
| Minimum | −11.76 | −11.09 | −8.94 | −12.42 |
| Maximum | 9.40 | 18.04 | 6.55 | 12.64 |
| Size | 25 | 30 | 25 | 30 |
| p-Value | 0.615 | 0.779 | ||
NMG, group of non-musicians; MG, group of musicians.
No statistically significant differences were observed in the comparison of the P300 amplitude suppression values between the NMG and the MG for the right ear [F1.53=0.257; p=0.615] and the left ear [F1.53=0.08; p=0.779].
Additionally, no statistically significant differences were observed in the comparison between the right and left ears for the P300 amplitude suppression values [NMG: t(24)=0.05; p=0.95; MG: t(29)=−0.814; p=0.42].
In the intra-group comparison, the P300 latency obtained in the absence and presence of contralateral noise was not significantly affected by the contralateral noise effect in the NMG for either the right ear [F(1,24)=1.446; p=0.241] or the left ear [F(1,24)=1.476; p=0.236]. However, in the MG, contralateral noise significantly affected the P300 latency value, causing a significant increase in this measure in both ears [right ear: F(1,29)=26.97; p<0.001; left ear: F(1,29)=9.861; p=0.004]. However, the P300 amplitude value was not significantly affected by the effect of contralateral noise in either group.
DiscussionThis study provided evidence that: (1) the MG had lower average latency values and higher average amplitudes compared to NMG in P300 without contralateral noise. This difference was statistically significant for amplitude values in both ears, but it was significant only in the right ear for latency values; (2) regarding P300 with contralateral noise, the MG also presented significantly higher amplitude values when compared with the NMG, although no statistically significant difference was observed between the groups for latency values; however, (3) no laterality effect was observed (difference between the right and left ears) in any of the groups; and (4) the analysis of the noise effect on the P300 response (intra-group analysis) demonstrated that the latency values were significantly increased in the MG. Nevertheless, this response was not observed in the NMG. In addition, for the amplitude values, the presence of noise did not significantly affect the response for either group.
Differences between musicians and non-musicians without contralateral stimulationThe superior performance reported in the MG, evidenced by lower latencies and higher amplitudes, corroborates the findings of other studies that observed lower latency for AEPs in musicians when compared with non-musicians.5,11 Although previous studies had verified differences between musicians and non-musicians in both ears,25–27 in the present study no statistically significant differences between latency values in left ear were observed. The authors attribute this fact to the small sample and the wide age range of the participants.
The P300 latency can be used as a measure of the speed of information processing in an oddball paradigm. Thus, it is suggested that the reduced latency observed in musicians may be related to the faster transmission and categorization of the stimulus, as well as the greater effectiveness in discriminating the target stimulus, which can be justified, in this case, by the musical stimulation.5
Regarding the P300 amplitude values, statistically significant differences were observed between the NMG and the MG for both right and left ears (Table 3). This finding is also in agreement with the literature, as higher amplitude values in musicians reflect more neural connections in the auditory pathway and suggest that musical training has an effect.3–5
It can be affirmed that musicians have enhanced auditory abilities compared with non-musicians. Several authors have associated greater auditory attentional capacity (higher concentration on the target stimulus), auditory memory, and the exceptional auditory perception of the musicians group with lower latency values and higher response amplitudes,28–30 as these individuals exhibit more accurate auditory information processing and more effective involving auditory pitch discrimination. Another possible cause of the lower latency values observed in the MG may be the presence of enhanced neural generators in the cortex, whose capacity to evaluate acoustic stimuli is better and faster than in non-musicians. Therefore, the stimulation analysis time is shorter in this group, resulting in smaller latencies.31
The improvement in auditory processing in musically trained individuals probably modifies the cortical organization, and those changes can be extended to sub-cortical sensory structures. Such changes have been observed throughout the auditory pathway, from the brain stem to the cortex.5 Certain authors also report that the brainstem responses of musicians are more robust, reflecting along the entire pathway. This greater robustness could be related to neural responses that are more synchronized to the onset of sound, which is characteristic of a highly functional peripheral auditory system.
How musicians perform on tasks that depend on auditory abilities motivates hypotheses concerning the impact of music training on brain mechanisms that underlie auditory attention and working memory. During such tasks (e.g., when subjects are instructed to listen for certain target tones or timbres), musicians demonstrate heightened recruitment of cortical areas associated with sustained auditory attention and working memory,17 such as the superior parietal cortex, as well as more consistent activation of prefrontal control regions.32 Indications that music training increases the contributions of the superior parietal and prefrontal cortices to active auditory processing and their roles in sustaining auditory attention and working memory may support the hypothesis that music training tunes the brain's auditory cognitive networks for cross-domain auditory processing.
Effect of contralateral noise on the responses of musicians and non-musiciansThis study demonstrated that P300 presented different behaviors in the presence and absence of contralateral noise, especially in musicians. The musician's average latency for P300 in the presence of contralateral noise was significantly later than without the contralateral noise. This difference in P300 latencies with and without contralateral noise was not observed in NMG. In addition, for the amplitude values, the presence of noise did not significantly affect the response for either group.
Studies of P300 with masking noise also demonstrated increased latencies with unaffected amplitudes.33,34 However, in a recent study, Schochat et al.22 did not observe differences regarding P300 latencies with and without contralateral noise in normal adults.
No statistically significant difference was observed between groups for P300 latency values with contralateral noise. Still, it is possible to note that MG latencies were more affected by contralateral noise in comparison to NMG, once MG latencies were later than NMGs.
However, for the P300 amplitude values, the difference between groups remained, and the MG presented significantly higher amplitude values compared with the NMG.
In addition, when comparing the suppression values between the groups, the musicians showed higher suppression values.
These findings, altogether, may be explained by the inhibitory effect the contralateral noise has on P300 responses, more specifically, on latency measures, which shows more vulnerability to this inhibitory effect. The latency measure has been related to the processing of auditory information and to auditory discrimination,35 and this is considered the most reliable measure in the P300 study.
Schochat el al.22 observed the presence of the inhibitory effect using a contralateral noise in both otoacoustic emission (OAE) and the late auditory potential P300. Studies using TOAE have described this inhibitory effect as the decrease of the cochlear response through the activation of the MOC in the presence of contralateral noise. This mechanism may provide an anti-masking effect that increases the discrimination of signal variation. In addition, it supplies a feedback gain-control system for moderate sounds levels that mediates selective attention and focuses attention during learning.36,37 Although the techniques used in the present study are different from those aforementioned, in general, the present findings corroborate the inhibitory effect actions, most probably by efferent system activation, using P300.
The function of the medial efferent system is complex and still not clear, as it involves different mechanisms of action mediated by the medial and lateral olivary tracts. However, evidence presented by several studies has shown that the MOCB plays an important role in the capacity of speech intelligibility in noise conditions. Recent studies using cochlear computer modeling and speech recognition have shown that the activation of the MOCB effect improved speech recognition in noise conditions.38 Thus, MOCB activity in humans should be of great importance for both the functioning of the peripheral auditory system and the improvement of auditory signal processing, especially for listening in noise conditions.39 The efferent system also contributes to the optimization of the discrimination of interaural differences at high frequency signals, increasing the difference between the information that reaches both cores of the superior olivary complex. This important role of the efferent pathway has repercussions on the ability to locate sounds of high frequency stimuli. Speech signals, especially consonants, are high frequency stimuli, and therefore, the integrity of the auditory system as a whole is required for verbal information to be appropriately processed.40
The authors also attribute the present findings to the action of the efferent system. Tomchik and Lu,41 in a study of animals’ auditory system, suggested that “primary afferent neurons adapt to noise, reducing their evoked firing rates in response to an additional stimulus, which may increase the latency of responses”. They also suggested that broadband noise disrupts the phase-locking of primary afferents to an added stimulus.
This line of reasoning leads to the belief that the effect of musical stimulation may constitute neurophysiologic evidence for the improvement of MOCB performance, maximizing the functioning of the afferent and efferent pathway, evidenced by the significant inhibitory effect found in musicians. In addition, it can be deduced that musicians show greater perceptual auditory advantages and enhanced speech processing, as well as advantages in frequency discrimination and temporal processing.
Another possible explanation for the present findings is related to the corticofugal modulation theory in humans. Studies have demonstrated that electrical stimulation in the auditory cortex may result in amplitude reduction of the OAE values in the contralateral ear. Other studies have also suggested a top-down control of the corticofugal descending auditory pathways in the medial olivocochlear efferent system.42,43 Other studies, who have failed to demonstrate correlations between OAE amplitude and suppression effect, also reinforce the hypothesis that there are other aspects influencing in MOC, such as cortical modulation.26,27
Even beyond sub-cortical and cortical functional enhancements, musical training may shape auditory function in structures that are as peripheral as the cochlea; musicians demonstrate a greater degree of efferent control over outer hair cell activity along the basilar membrane than non-musicians.15 Such comprehensive perceptual and neural enhancements may be driven, at least in part, by strengthened cognitive control over basic auditory processing, as engendered by auditory attention32,44 and working memory,17,18 two auditory cognitive skills that are enhanced in musicians.
Thus, it can be stated that musical training may be an important tool for auditory training, prevention, habilitation, and reversion of a wide range of auditory processing deficits.
Regarding the absence of the laterality effect in both groups and both evaluated conditions, the authors believe that these results can be explained by the fact that both hemispheres participate in this cognitive processing task. This supports the view that P300 reflects an integration of function of various areas of the brain.
Some shortcomings of this study may have influenced the results: absence of a sample size calculation, broad age range, broad range of years of formal music practice, the absence of manual dominance as an inclusion criterion, and the long duration of the test. Nevertheless, the authors believe that this study presents promising results regarding the use of new tolls to verify the medial olivocochlear efferent system activity, as well as new information on this activity through a LLAEP in musicians. Future studies should take these shortcomings into consideration.
ConclusionIn this study, evidence about cortical inhibition effect was found using an ERP (P300) in the presence and absence of contralateral noise condition in both groups. Besides, musicians demonstrated a greater inhibition effect in comparison with non-musicians, evidencing that the central auditory nervous system of the former presents characteristic peculiarities due to the musical practice to which they are constantly exposed.
Conflicts of interestThe authors declare to have no conflicts of interest.
Please cite this article as: Rabelo CM, Neves-Lobo IF, Rocha-Muniz CN, Ubiali T, Schochat E. Cortical inhibition effect in musicians and non-musicians using P300 with and without contralateral stimulation. Braz J Otorhinolaryngol. 2015;81:63–70.
Institution: Departament of Physiotherapy, Speech and Language Therapy, Faculdade de Medicina, Universidade de São Paulo (FMUSP), São Paulo, SP, Brazil.
gology is pleased to honor the reviewers


