

As funções atribuídas aos andrógenos aumentaram, variam desde o papel no eixo hipotálamo‐hipófise‐gonadal e comportamentos reprodutivos até a modulação da cognição, humor e outras funções. As diferenças entre os sexos e as mudanças nos hormônios sexuais circulantes afetam a função sensorial humana. Na literatura, os autores relataram esse tipo de influência para o olfato, principalmente no sexo feminino.
ObjetivoInvestigar os efeitos dos baixos níveis de testosterona nas funções olfativas em homens, neste estudo clínico prospectivo.
MétodoPacientes do sexo masculino com diagnóstico de câncer de próstata foram incluídos no estudo. Compreenderam o grupo de estudo 39 pacientes com câncer de próstata cujos níveis de testosterona eram inferiores a 50 ng/dL devido à castração. Foram determinados como grupo controle 31 pacientes com câncer de próstata que não foram emasculados, com níveis de testosterona superiores a 50 ng/dL. Testes de rinometria acústica e pico de fluxo inspiratório nasal foram feitos para todos os participantes; e para avaliação da função olfativa, ambos os grupos concluíram o teste olfativo do Connecticut chemosensory clinical research center.
ResultadosA média da idade dos pacientes e controles foi de 69,6±7,2 (57±89) e 66,3±5,8 (50±78) anos, respectivamente (p=0,039). Houve uma diferença significante entre os grupos em relação a níveis de testosterona (p <0,0001). A regressão logística multivariada revelou o nível de testosterona como o único fator preditivo que determinou a diferença entre os grupos. Em termos de parâmetros olfativos, todos os escores foram menores no grupo castrado (teste do limiar de butanol p=0,019, identificação p=0,059 e escore do Connecticut centerp=0,029). Houve uma correlação significante entre o nível de testosterona e os parâmetros olfativos (p=0,023; p=0,025 para identificação e escore do Connecticut center, respectivamente).
ConclusãoBaixos níveis de testosterona em homens têm efeito negativo na função olfativa. Mais pesquisas moleculares são necessárias para entender a conexão entre testosterona e olfação.
O sentido do olfato é parte de um sistema neurossensorial que desempenha papéis importantes na vida humana. Os distúrbios olfativos podem afetar negativamente a qualidade de vida, a segurança e a nutrição. Qualquer doença que afete as regiões localizadas na via anatômica do sistema olfativo, da cavidade nasal até o sistema nervoso central (SNC), pode estar envolvida na etiologia dos distúrbios olfativos. Além dessas alterações anatômicas, algumas condições endocrinológicas, como a doença de Addison, a síndrome de Turner ou o hipotireoidismo, podem causar esses distúrbios.1
As diferenças entre os sexos e mudanças no ciclo menstrual afetam a função sensorial humana, mesmo que não causem distúrbios olfativos. A associação entre a função olfativa humana e os hormônios reprodutivos é complexa; e uma relação simples entre níveis de hormônios gonadais circulantes e olfação raramente está presente.2
Alguns estudos demonstram que as funções olfativas são alteradas por mecanismos centrais e periféricos que se correlacionam com alterações no nível dos hormônios esteroides em mulheres.2–5 Entretanto, o mecanismo exato que modula a percepção olfativa é desconhecido. Com relação à testosterona e sua associação com o olfato, encontramos menos estudos relevantes na literatura, quase todos relacionados a animais.
As funções atribuídas aos andrógenos aumentaram nos últimos anos, desde desempenhar um papel bem conhecido no eixo hipotálamo‐hipófise‐gonadal e comportamentos reprodutivos até a modulação de funções como cognição e humor. Os efeitos da testosterona no sistema nervoso são mediados por receptores de andrógenos ou estrógenos (após aromatização neural).6,7
O hipogonadismo é caracterizado por baixos níveis de testosterona, os quais causam impotência, perda da libido, sintomas vasoativos, dislipidemia e ganho de peso; dessa forma, níveis reduzidos de testosterona afetam muitos sistemas orgânicos.8
O câncer de próstata (CP) é um dos tipos de câncer mais comuns nos homens. Em 1941, Huggins relatou a proliferação e o crescimento do epitélio da próstata; desde então, a terapia de privação androgênica (ADT, do inglês Androgen Deprivation Therapy) estabeleceu sua posição como a principal opção de manejo de CP avançado.9,10
A ADT melhora a taxa de sobrevida e a qualidade de vida desses pacientes. Os agonistas do hormônio liberador de gonadotrofina (GnRH) e antiandrogênicos são a terapia hormonal inicial mais usada. A ADT causa hipogonadismo em pacientes com CP.11
As diferenças entre os sexos e as mudanças nos hormônios sexuais circulantes afetam a função sensorial humana. Na literatura, os autores relataram esse tipo de influência para a audição, visão, gustação e olfação. Que seja de nosso conhecimento, somos os primeiros a demonstrar a relação entre os níveis de testosterona e a olfação em homens.
Neste estudo, pretendemos demonstrar como os baixos níveis de testosterona causados pelo ADT afetam a função olfativa, juntamente com alguns parâmetros nasais.
MétodoA aprovação do comitê de ética foi obtida de nosso conselho de ética institucional (data: 7 de dezembro de 2018, número de aprovação: 1556). Depois que todos os participantes incluídos no estudo terem sido informados sobre os testes de função nasal, eles deram seu consentimento informado para participar. Este estudo prospectivo e controlado foi feito entre junho de 2018 e fevereiro de 2019. Foram incluídos pacientes do sexo masculino com diagnóstico de CP, divididos em dois grupos. Entre nossa amostra, 39 pacientes com CP cujos níveis de testosterona eram inferiores a 50 ng/dL devido à castração cirúrgica ou médica feita na clínica de urologia do nosso hospital constituíram o grupo de estudo, enquanto 31 pacientes não emasculados com CP cujos níveis de testosterona eram superiores a 50 ng/dL foram considerados como o grupo controle.
Todos os possíveis participantes foram examinados através de endoscopia nasal e foram excluídos do estudo indivíduos com s nasosinusais que pudessem alterar a sensação olfativa, como desvio de septo, polipose nasal ou sinusite aguda. Pacientes submetidos à cirurgia nasal ou com histórico de traumatismo craniano e infecção do trato respiratório superior identificado durante o exame foram excluídos.
Para obter medidas objetivas, os testes de rinometria acústica (RA) (RhinoScan, Manual v. 2.6 ed. 1.1, RhinoMetrics®, Dinamarca) e pico de fluxo inspiratório nasal (PFIN) foram feitos antes e após o uso de spray nasal descongestionante (cloridrato de xilometazolina, 0,01%). Para avaliação da função olfativa, todos os participantes concluíram o teste olfativo do Connecticut chemosensory clinical research center (CCCRC).
A idade dos indivíduos, o nível de testosterona, o índice de massa corporal, o histórico de tabagismo, as doenças subjacentes, os valores mínimos da área de seção transversal (MCA) 1 e 2 em RA e o PFIN, limiar de butanol, identificação de odor e escores do teste CCCRC foram comparados entre os dois grupos.
Avaliação da função olfativaPara o teste do limiar de butanol, as soluções diluídas de butanol foram preparadas e alinhadas de 0 a 9, onde 0 representa a solução mais concentrada e 9 representa a solução mais diluída. O concentrado mais forte (frasco 0) foi butanol a 4% em uma solução de água destilada. Cada diluição subsequente (frascos 1 a 9) consistiu em uma proporção de 1:3 de butanol para água destilada. Foram apresentados aos pacientes dois frascos de aparência idêntica, embora um contivesse água destilada e o outro o concentrado de butanol diluído. Os indivíduos foram instruídos a ocluir uma narina e colocar a ponta do primeiro frasco 3cm abaixo da outra narina. Uma vez que cada indivíduo identificasse corretamente a mesma concentração de butanol cinco vezes consecutivas, a pontuação era registrada para essa narina. A outra narina foi então testada separadamente e as pontuações para ambas as narinas foram calculadas como média para atingir a pontuação final. Os escores possíveis variaram de 0 a 9 pontos, mas todas os escores de 7 pontos ou mais foram registrados como 7 pelo teste do CCCRC.
Como teste de identificação, os odores do teste do CCCRC que Veyseller et al.12 consideraram apropriado para uso na população turca foram usados. Vick Vaporub®, sabão, manteiga de amendoim, bolas de naftalina, cacau, café, canela e talco de bebê foram apresentados aos indivíduos em frascos opacos. Eles receberam uma lista de múltipla escolha, com os seguintes itens de distração: papel queimado, Vick Vaporub®, cacau, talco de bebê, manteiga de amendoim, café, menta, canela, sabão, bolas de naftalina, geleia, ketchup, pimenta e plástico. A capacidade de sentir o cheiro de Vick Vaporub® indicava uma função intacta do nervo trigêmeo; esse item foi facilmente identificado por todos os indivíduos e não foi incluído no escore final. Os escores para as duas narinas foram calculados e todos os escores que variaram de 0 a 7 foram anotados.
O limiar de butanol e escores de identificação foram calculados como média para uma pontuação final composta do CCCRC para determinar a capacidade olfativa de cada indivíduo.
Análise estatísticaOs resultados foram analisados estatisticamente com o softwareStatistical Package for Social Sciences, versão 18. A adequação da distribuição normal de cada variável foi analisada através de métodos visuais (histograma e gráficos de probabilidade) e analíticos (testes de Kolmogorov‐Smirnov e Shapiro‐Wilk). A estatística descritiva foi dada como média e desvio‐padrão para variáveis numéricas normalmente distribuídas e como mediana e intervalo interquartil para variáveis numéricas distribuídas de forma não normal. Um teste t independente foi implaentado para comparar duas variáveis independentes, enquanto o teste U de Mann‐Whitney foi aplicado para variáveis que não apresentaram distribuição normal. Os coeficientes de correlação e significância estatística para relações entre variáveis numéricas distribuídas de forma anormal foram medidos pelo teste de correlação de Spearman, onde variáveis categóricas foram expressas em números e porcentagens. As diferenças entre os grupos foram comparadas com o teste do qui‐quadrado e o teste exato de Fisher. Uma análise de regressão logística multivariada foi feita para determinar o efeito independente de cada variável na variável de desfecho. O nível de significância estatística foi estabelecido como <0,05.
ResultadosForam analisados 39 pacientes do grupo 1 com baixo nível de testosterona e 31 pacientes não castrados do grupo 2 (grupo controle), entre os quais todos foram diagnosticados com CP. No grupo controle, 19 pacientes foram submetidos a prostatectomia radical, 4 pacientes a radioterapia e 8 pacientes foram submetidos a vigilância ativa. No grupo com baixo nível de testosterona, 37 pacientes foram emasculados clinicamente (como análogos do hormônio liberador do hormônio luteinizante, 32 pacientes receberam acetato de leuprolide; 5 pacientes receberam acetato de goserelina), enquanto 2 pacientes foram emasculados cirurgicamente (orquiectomia bilateral).
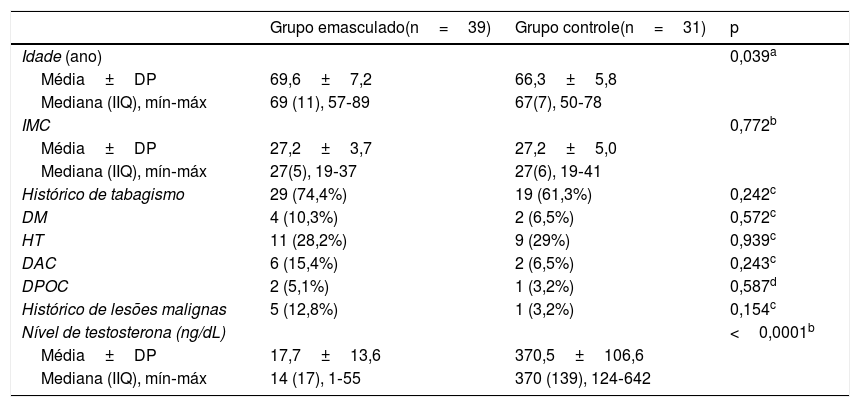
A média da idade dos pacientes do grupo 1 e do grupo 2 foi de 69,6±7,2 (57±89) e 66,3±5,8 (50±78) anos, respectivamente (p=0,039). Uma diferença significante foi identificada entre os grupos em termos de níveis de testosterona (p <0,0001), embora nenhuma diferença estatisticamente significante tenha sido detectada entre os dois grupos em termos de outras características demográficas (tabela 1). A regressão logística multivariada revelou o nível de testosterona como o único fator preditivo que determinou a diferença entre os grupos.
Comparação das características demográficas dos voluntários
| Grupo emasculado(n=39) | Grupo controle(n=31) | p | |
|---|---|---|---|
| Idade (ano) | 0,039a | ||
| Média±DP | 69,6±7,2 | 66,3±5,8 | |
| Mediana (IIQ), mín‐máx | 69 (11), 57‐89 | 67(7), 50‐78 | |
| IMC | 0,772b | ||
| Média±DP | 27,2±3,7 | 27,2±5,0 | |
| Mediana (IIQ), mín‐máx | 27(5), 19‐37 | 27(6), 19‐41 | |
| Histórico de tabagismo | 29 (74,4%) | 19 (61,3%) | 0,242c |
| DM | 4 (10,3%) | 2 (6,5%) | 0,572c |
| HT | 11 (28,2%) | 9 (29%) | 0,939c |
| DAC | 6 (15,4%) | 2 (6,5%) | 0,243c |
| DPOC | 2 (5,1%) | 1 (3,2%) | 0,587d |
| Histórico de lesões malignas | 5 (12,8%) | 1 (3,2%) | 0,154c |
| Nível de testosterona (ng/dL) | <0,0001b | ||
| Média±DP | 17,7±13,6 | 370,5±106,6 | |
| Mediana (IIQ), mín‐máx | 14 (17), 1‐55 | 370 (139), 124‐642 |
IIQ, Intervalo interquartil; IMC, Índice de massa corporal; DM, Diabetes mellitus; HT, Hipertensão; DAC, Doença arterial coronariana; DPOC, Doença pulmonar obstrutiva crônica.
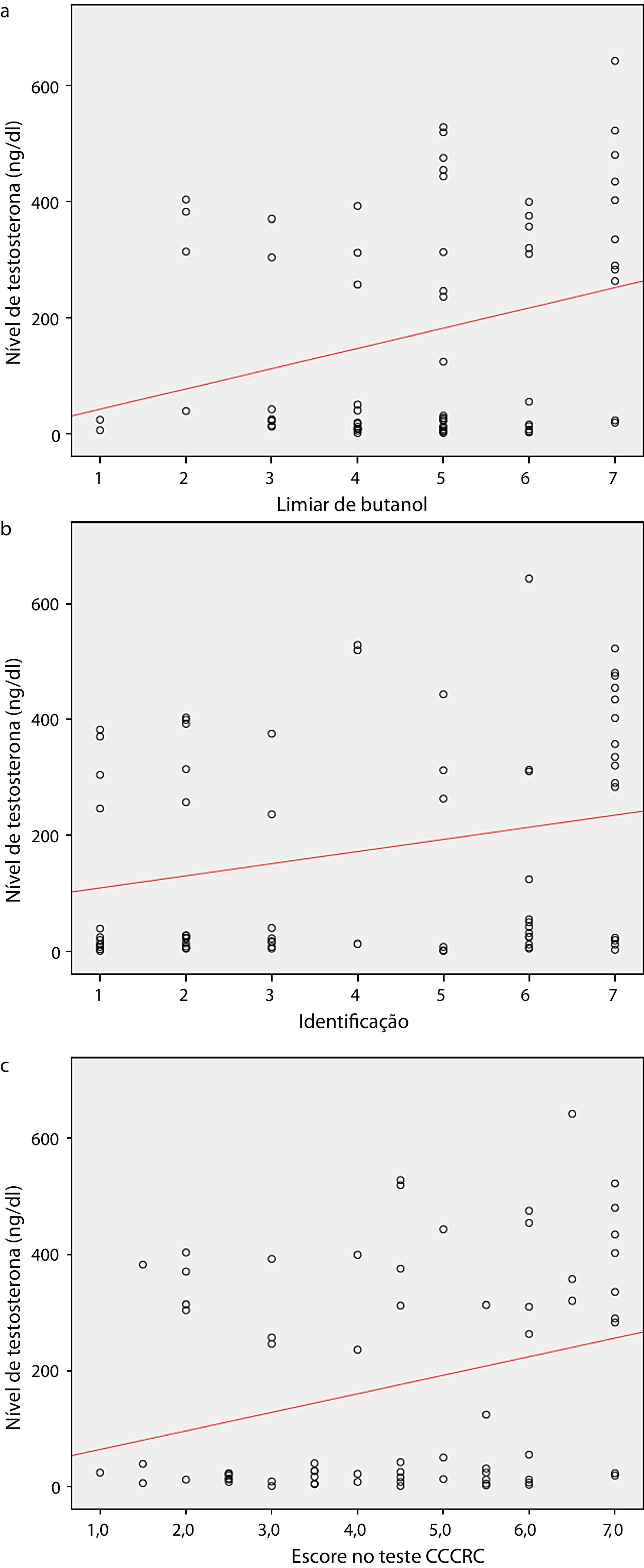
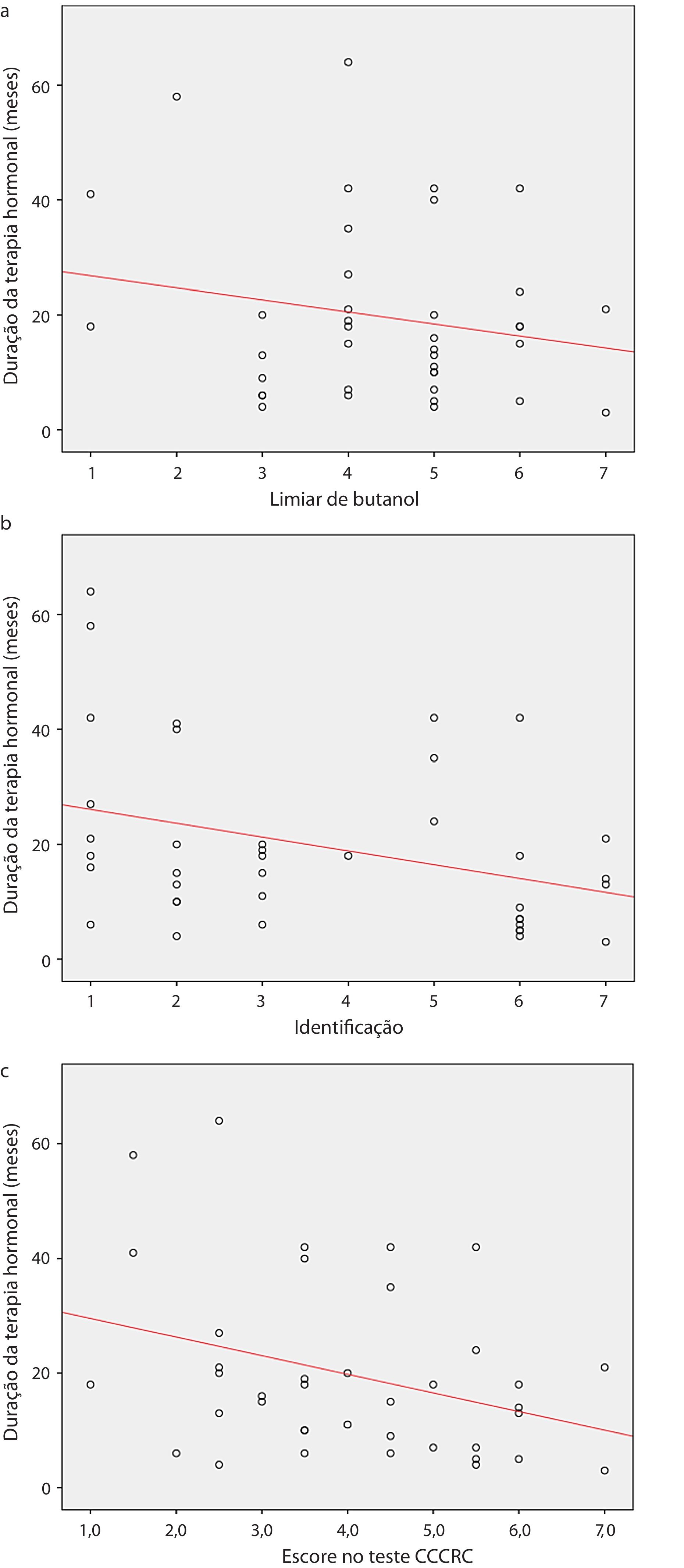
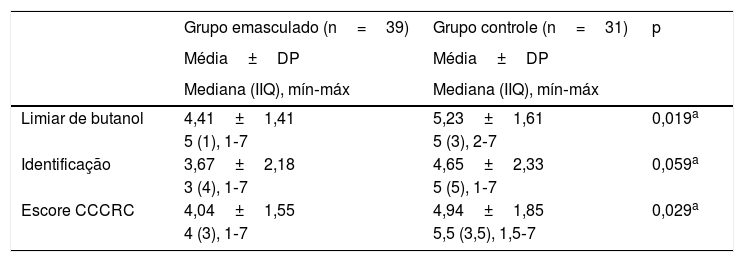
Quando os dois grupos foram comparados em termos de parâmetros olfativos, todos os escores foram menores no grupo castrado (teste do limiar de butanol p=0,019, teste de identificação p=0,059 e escore do CCCRC p=0,029) (tabela 2). Foi identificada uma correlação significante entre o nível de testosterona e os parâmetros olfativos (para teste de identificação: p=0,023 e para escore no CCCRC: p=0,025) (fig. 1a, b and c). Além disso, no grupo de baixa testosterona, uma correlação significantemente negativa foi determinada exclusivamente entre a duração da terapia hormonal e os parâmetros olfativos em termos de identificação (p=0,017) (fig. 2a, b, and c). No grupo castrado, os escores de identificação e CCCRC foram menores entre os pacientes cuja duração da terapia hormonal foi superior a 9 meses (tabela 3).
Comparação de grupos a partir do ponto de olfação
| Grupo emasculado (n=39) | Grupo controle (n=31) | p | |
|---|---|---|---|
| Média±DP | Média±DP | ||
| Mediana (IIQ), mín‐máx | Mediana (IIQ), mín‐máx | ||
| Limiar de butanol | 4,41±1,41 | 5,23±1,61 | 0,019a |
| 5 (1), 1‐7 | 5 (3), 2‐7 | ||
| Identificação | 3,67±2,18 | 4,65±2,33 | 0,059a |
| 3 (4), 1‐7 | 5 (5), 1‐7 | ||
| Escore CCCRC | 4,04±1,55 | 4,94±1,85 | 0,029a |
| 4 (3), 1‐7 | 5,5 (3,5), 1,5‐7 |
Escore CCCRC, teste de olfato do Connecticut chemosensory clinical research center; IIQ, Intervalo interquartil.
 Nível de testosterona e limiar de butanol; (b) nível de testosterona e identificação de odor; (c) Nível de testosterona e escore no CCCRC, teste de olfato do Connecticut chemosensory clinical research center.")
 Duração da terapia hormonal e limiar de butanol; (b) Duração da terapia hormonal e identificação de odor; (c) Duração da terapia hormonal e escore no CCCRC, teste de olfato do Connecticut chemosensory clinical research center.")
Relação entre a duração da terapia hormonal e parâmetros olfativos
| Duração da TH ≤ 9 meses(n=11) | Duração da TH> 9meses(n=28) | p | |
|---|---|---|---|
| Limiar de butanol | 0,648a | ||
| ≤ 4 | 6 (54,5%) | 13 (46,4%) | |
| > 4 | 5 (45,5%) | 15 (53,6%) | |
| Identificação | 0,012a | ||
| ≤ 4 | 3 (27,3%) | 20 (71,4%) | |
| > 4 | 8 (72,7%) | 8 (28,6%) | |
| Escore CCCRC | 0,037a | ||
| ≤ 4 | 3 (27,3%) | 18 (64,3%) | |
| > 4 | 8 (72,7%) | 10 (35,7%) |
TH, Terapia hormonal; Escore CCCRC, teste de olfato do Connecticut chemosensory clinical research center.
Em nosso estudo, verificamos que a função olfativa é pior em pacientes com CP cujos níveis de testosterona são inferiores a 50 ng/dL do que em pacientes com CP que apresentam níveis de testosterona superiores a 50 ng/dL. Esse achado implica que a testosterona pode ter influência na função olfativa, embora possa surgir a questão de como a testosterona e a olfação estão conectadas.
A perda olfativa pode resultar de um distúrbio de condução que impede que os odorantes inspirados atinjam a fenda olfativa do nariz, ou um distúrbio neurossensorial nos neurônios do receptor olfativo (RO) ou em suas projeções centrais.
Na literatura, observamos hipogonadismo com disfunção olfativa. Na síndrome de Kallmann (SK) – também conhecida como hipogonadismo hipogonadotrófico congênito – o hipogonadismo ocorre com anosmia.13
Em SK, as mutações genéticas causam falha na migração dos neurônios GnRH dos placódios olfatórios para seu destino hipotalâmico e lobo olfativo e distúrbio neuronal olfativo.14 O GnRH e os neurônios olfativos compartilham origens embriológicas do desenvolvimento,15 pois ambos se originam de células‐tronco dentro do placódio olfatório embrionário.16 Os neurônios GnRH migram para o hipotálamo através dos nervos vomeronasais.17,18 Aqui, observamos o GnRH e a região olfativa conjuntamente.
O ciclo de feedback endócrino clássico ocorre no sistema reprodutor masculino. O GnRH hipotalâmico estimula o hormônio luteinizante da hipófise (LH) e o hormônio folículo estimulante (FSH), ambos estimulam os testículos. O LH estimula a secreção de testosterona pelas células de Leydig. Após a castração, pode haver alterações no mecanismo de feedback e no ambiente neuroendócrino. A relação entre olfação e testosterona pode ocorrer via GnRH no arco. Embora o GnRH e os receptores de esteroides estejam presentes nas regiões de processamento olfativo dos vertebrados, os níveis de mRNA do receptor de andrógeno no bulbo olfativo de peixes da família Cichlidae foram considerados positivamente correlacionados com os níveis de androgênio circulantes.19 No cérebro adulto, essas células também são encontradas nos nervos/bulbo olfativos e núcleos olfativos mediais.20 As células do GnRH também são encontradas em locais ectópicos, como a submucosa nasal e o tecido conjuntivo entre o epitélio olfativo (EO) e o bulbo olfativo em camundongos.21
O nervus terminalis (NT) é um nervo organizado contendo GnRH que se estende da cavidade nasal às regiões do cérebro, media o processamento e a reprodução quimiosensorial.22,23 Wirsig‐Wieshmann et al. relataram a expressão do receptor de GnRH na mucosa olfativa e sugeriram que o GnRH modula a atividade das células associadas à quimiorrecepção, que é o potencial de alterar a olfação.24
Kawai et al. sugeriram que em peixinhos dourados, o NT desempenha papel crítico, media a capacidade de resposta olfativa dos animais.25
Neuhaus et al. demonstraram que o status do receptor de esteroide acoplado à proteína G (PSGR) específico da próstata nas células da próstata é um componente ectópico da superfamília RO e implica funcionalidade, porque pode ser ativado por ligantes. Os transcritos de PSGR também podem ser distinguidos no EO humano.26,27
As relações entre os fatores neuroendócrinos e o sistema olfativo humano são muito complexas. A possível conexão pode não ser simples como relação direta entre olfação e níveis hormonais gonadais circulantes de maneira síncrona; em vez disso, podem existir muitos mecanismos possíveis. Se essa associação não for via permeabilidade nasal, fluxo aéreo ou alterações do muco olfatório, então essas possibilidades podem incluir uma influência em sistemas inespecíficos de excitação cerebral, como o sistema de ativação reticular, efeito direto nas vias de transdução olfativa do SNC ou influência indireta de outros sistemas endócrinos nas vias olfativas do SNC.2,28
Outra conexão possível pode ser um efeito direto do hormônio testosterona na olfação. Sawa Horie et al., sugeriram síntese de novo e/ou metabolismo de esteróides sexuais em células da mucosa olfativa em ratos.29 Os hormônios gonadais passam para o cérebro e os estudos sobre os hormônios esteroides marcados sugerem que a retenção hormonal no cérebro tem especificidade regional. Estrógenos e andrógenos se ligam a núcleos no bulbo olfativo5,30–32 e a enzima aromatase, necessária para a conversão de andrógeno em estrogênio, também é expressa no bulbo olfativo.33,34 Kass et al. sugeriram que os esteroides sexuais de alguma forma modulam a atividade dos neurônios sensoriais primários no principal EO.35 O cérebro humano é sensível às ações dos esteróides gonadais; em algumas estruturas olfativas, ele contém receptores para andrógenos e estrógenos.36,37
Foi relatada uma sensibilidade olfativa alterada que se correlaciona com as alterações nos níveis hormonais dos esteroides reprodutivos no sexo feminino,2–5 principalmente por seus efeitos positivos na função olfativa; entretanto, para a testosterona, alguns estudos na literatura sugeriram a ocorrência de efeitos negativos. Kanageswaran et al. relataram os efeitos moduladores da progesterona e do estradiol nas respostas evocadas por odorantes nos neurônios do RO em camundongos.38 Kass et al. sugeriram que os hormônios gonadais podem facilitar a detecção de odores e a discriminação de odorantes semelhantes em mulheres, embora reduzam essa detecção em homens.35 Pietras e Moulton5 relataram que a testosterona em doses suprafisiológicas melhorou a capacidade de detecção de odor em ratos fêmeas gonadectomizadas (cuja aromatização da testosterona para estrógenos pode ser um mecanismo possível), mas a castração masculina não influencia a sensibilidade dos ratos machos à urina no estro feminino ou ao acetato de etila.2,39,40
Fizemos o presente estudo para avaliar mais detalhadamente o possível papel que a testosterona tem em homens e identificamos diminuição da função olfativa (ou seja, pior limiar de butanol e pior identificação de odores) em homens com baixos níveis de testosterona. Identificamos ainda uma correlação significante entre os níveis de testosterona e os parâmetros olfativos.
A literatura indica que camundongos machos e fêmeas que não têm hormônios gonadais circulantes não conseguem fazer a tarefa Go/No‐Go.41,42 Kunkhyen et al.42 relataram que a administração de propionato de testosterona em camundongos machos castrados traz a capacidade de discriminação do odor urinário de volta aos níveis pré‐gonadectomia; hormônios gonadais, mas não o sexo, afetam a aquisição e manutenção de uma tarefa de discriminação de odores Go/No‐Go. Doty e Ferguson‐Segall40 indicaram que a castração influencia a capacidade do rato macho de melhorar sua capacidade de detecção de odores ao longo do tempo.
Outra conexão possível entre a testosterona e a função olfativa pode ser a função cognitiva mediada pelos hormônios gonadais. O nível de testosterona biodisponível está associado à função cognitiva em homens idosos.43 A suplementação de testosterona melhora a memória de trabalho em homens idosos que têm níveis reduzidos de testosterona livre.44 Em ratos machos, Gibbs et al.45 relataram que a testosterona tem influência significativa em domínios cognitivos específicos.
Também examinamos as vias aéreas nasais para detectar os problemas de condução que podem ser responsáveis pela disfunção olfativa. Os testes feitos aqui (RA e PFIN) forneceram dados para fazer uma comparação entre os níveis de testosterona. Os valores de MCA 1‐2 na RA e PFIN não revelaram diferenças significantes entre os dois grupos, o que aponta para um mecanismo neurossensorial mais possível da disfunção olfativa.
Quase todos os estudos em animais e humanos que dizem respeito à influência do sexo e dos hormônios sexuais circulantes na principal função olfativa avaliaram os limiares de detecção de odorantes. Além disso, estudamos a capacidade de nossos indivíduos de identificar odores e determinamos que o grupo com baixa testosterona alcançou resultados significantemente piores do que o grupo não castrado no teste de limiar de butanol. A identificação de odores também foi pior com níveis baixos de testosterona. Verificou‐se que o nível de testosterona está significantemente correlacionado com a capacidade de identificação olfativa.
A duração do tempo com baixos níveis de testosterona é importante para o desenvolvimento de disfunção olfativa dos pacientes. Determinamos uma correlação significantemente negativa entre a duração da ADT e a identificação de odores.
Não foi possível determinar um mecanismo exato de associação entre o hormônio testosterona e a disfunção olfativa em estudos em animais e em laboratório publicados anteriormente. Que seja de nosso conhecimento, este é o primeiro estudo em humanos que demonstra os efeitos da testosterona na olfação em homens. Este estudo também revelou alguns resultados definitivos e correlações relacionadas. Isso não significa que “sem testosterona, não há olfação”, embora uma associação pareça ser possível.
Um estudo pode ser elaborado com um grupo de pacientes com níveis reduzidos de testosterona, cujos testes de olfação devem ser feitos antes e após a reposição do hormônio testosterona. Mas esse projeto não é possível para esses grupos de pacientes. Assim, para obter resultados mais definitivos, a avaliação e comparação olfativa devem ser feitas antes e depois da castração terapêutica de cada paciente.
ConclusãoEste estudo demonstrou que o nível de testosterona pode ter efeitos na função olfativa em homens. Nos homens, o baixo nível de testosterona afeta negativamente o limiar olfativo e a identificação de odorantes voláteis. O aumento do tempo com baixa testosterona agrava a identificação do odor. Mais pesquisas moleculares são necessárias para entender o mecanismo real e exato que estabelece a conexão entre a testosterona e o olfato.
Aprovação éticaTodos os procedimentos feitos em estudos que envolvem participantes humanos ocorreram em conformidade com os padrões éticos do comitê de pesquisa institucional (Istanbul Training and Research Hospital's Ethics Committee; número de referência: 1556) e com a declaração de Helsinque de 1964 e suas alterações posteriores ou padrões éticos comparáveis.
Consentimento informadoO consentimento informado foi obtido de todos os participantes incluídos no estudo.
Conflitos de interesseOs autores declaram não haver conflitos de interesse.
FinanciamentoOs autores não receberam apoio financeiro para a pesquisa, autoria e publicação deste artigo. Este manuscrito foi apresentado no 15° Congresso de Rinologia Turca, de 4 a 7 de abril de 2019, em Antalya, Turquia.
Conflitos de interesseOs autores declararam não haver conflitos de interesse.
Como citar este artigo: Kırgezen T, Yücetaş U, Server EA, Övünç O, Yiğit Ö. Possible effects of low testosterone levels on olfactory function in males. Braz J Otorhinolaryngol. 2021;87:702–10.
A revisão por pares é da responsabilidade da Associação Brasileira de Otorrinolaringologia e Cirurgia Cérvico‐Facial.
gology tem o prazer em homenagear os revisores


